
Descripción científica de la enfermedad del tizón temprano (2020)
Basado en Adolf B. et al. (2020) Enfermedades fúngicas, oomicetas y plasmodioforidas de la papa.
Los autores de este contenido son Hans Hausladen y Birgit Adolf
Síntomas
Los síntomas foliares del tizón temprano (EB, por sus siglas en inglés) son necrosis de color marrón oscuro a negro. Los primeros síntomas foliares se vuelven visibles en las hojas más bajas y, por lo tanto, más viejas, apenas unas semanas después de la emergencia. Inicialmente aparecen manchas puntiformes de color marrón oscuro, de unos pocos milímetros de diámetro. El área necrótica aumenta gradualmente y los síntomas en la hoja crecen hasta ocupar todo el tejido verde foliar (imagen inferior).
A menudo, las lesiones quedan restringidas por las nervaduras de la hoja y adquieren una forma angular. El tamaño de la necrosis puede variar en ancho, desde unos pocos milímetros hasta 2 cm. Dentro de las lesiones más grandes, puede observarse una serie de anillos concéntricos oscuros.
Este patrón en forma de diana es típico de los síntomas del tizón temprano. Posteriormente, el tejido foliar necrótico suele quedar rodeado por un borde clorótico causado por micotoxinas fúngicas, que vuelven amarillo el tejido de la hoja. La clorosis puede extenderse a toda la hoja infectada. Durante el avance del tizón temprano, las áreas infectadas se agrandan, toda la hoja se vuelve necrótica y cae.
En Europa, se produce un fuerte aumento de la infestación de tizón temprano desde mediados de julio en adelante, especialmente durante períodos cálidos y secos o cuando el cultivo de papa se encuentra bajo estrés, y en plantas fisiológicamente más viejas. Luego, el tizón temprano comienza a propagarse desde las hojas inferiores hacia los niveles medios y finalmente superiores del follaje.

Síntomas iniciales (izquierda) y avanzados (derecha) del tizón temprano de la papa en el follaje
Los conidios de Alternaria que son lavados de las hojas también pueden infectar los tubérculos. Los síntomas del tizón temprano en los tubérculos son lesiones oscuras, ligeramente hundidas (imagen inferior). La pudrición seca o dura de los tubérculos causa pérdidas durante el almacenamiento, reduce la calidad de las papas de mesa y disminuye la capacidad de germinación de las papas semilla.

Síntomas del tizón temprano de la papa en tubérculos
No es posible distinguir entre las diferentes especies de Alternaria que causan el tizón temprano basándose únicamente en los síntomas, aunque a veces los síntomas denominados mancha marrón de la hoja —manchas pequeñas, irregulares a circulares, de color marrón oscuro, cuyo tamaño va desde un punto hasta 4 mm— se atribuyen a A. alternata (Fairchild et al. 2013).
Organismo causal
El principal agente causal del tizón temprano en cultivos solanáceos se considera generalmente Alternaria solani Sorauer (Gannibal et al. 2014). Sin embargo, existen informes de otras especies de Alternaria de esporas grandes involucradas en el tizón temprano de la papa.
Rodrigues et al. (2010) encontraron que A. grandis Simmons, y no A. solani, era el agente causal del tizón temprano que afectaba a plantas de papa en varias regiones productoras de Brasil, y Duarte et al. (2014) confirmaron que esta especie puede causar tizón temprano en papa en ensayos de campo mediante inoculación artificial con A. grandis.
Alternaria protenta ha sido detectada como la especie causal de Alternaria para el tizón temprano en Argelia (Ayad et al. 2017) y, junto con A. grandis y A. solani, fue encontrada como parte del complejo de especies de Alternaria detectadas en lesiones de tizón temprano en Bélgica (Landschoot et al. 2017a).
Entre las especies de Alternaria de esporas pequeñas, A. alternata aparece regularmente en lesiones de tizón temprano, pero se considera un invasor secundario (Leiminger et al. 2014; Stammler et al. 2014; Rotem 1994).
Alternaria solani sobrevive el invierno como micelio, clamidosporas o conidios en el suelo y en restos vegetales infestados (ver ciclo de la enfermedad en la imagen inferior). En primavera, la infección primaria ocurre a través del inóculo (conidios) transportado a las hojas inferiores por salpicaduras de lluvia. El patógeno puede penetrar el tejido foliar directamente a través de la epidermis intacta o por estomas y heridas.
Los primeros síntomas del tizón temprano en las hojas se vuelven visibles entre 2 y 4 semanas después de la emergencia del cultivo de papa. Inicialmente, se infestan las hojas más viejas cercanas al suelo. El hongo queda restringido al nivel inferior del follaje durante varias semanas y al principio parece no ser motivo de preocupación.
La formación de conidios ocurre en el tejido foliar necrótico a temperaturas entre 5 y 30 °C (óptimo 20 °C). Este inóculo secundario se disemina por el viento y causa infecciones en las hojas y tallos circundantes. El período de latencia promedia entre 3 y 7 días. Bajo condiciones favorables para la infección, y una vez que la planta alcanza cierta edad, A. solani coloniza muy rápidamente los niveles medios y superiores del follaje.
En el campo, puede observarse una progresión en cascada del hongo desde las hojas inferiores, pasando por las medias, hasta las superiores. Las hojas fuertemente infestadas caen y permanecen como fuente de inóculo sobre y dentro del suelo.
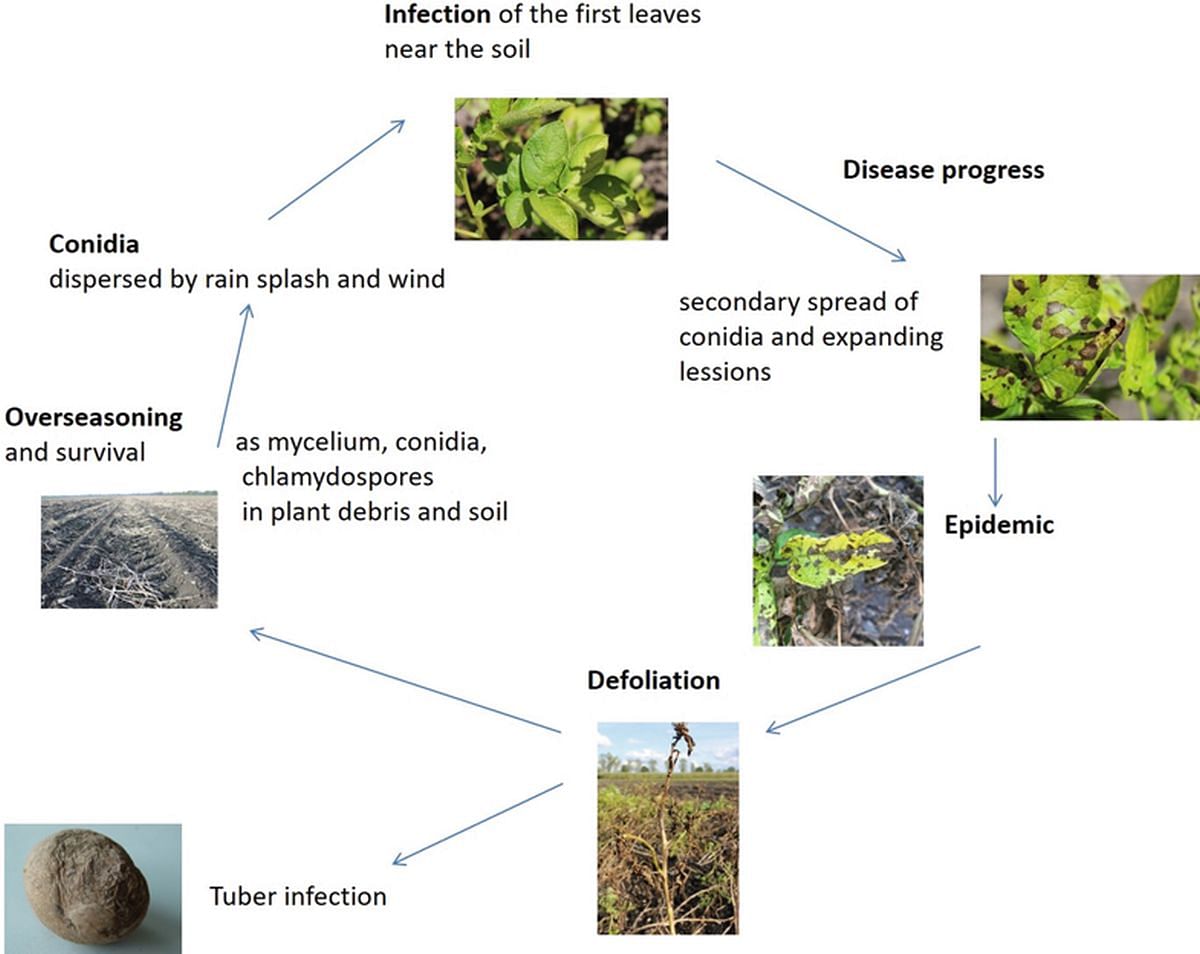
Ciclo de la enfermedad del tizón temprano de la papa
La progresión del tizón temprano depende de parámetros climáticos, la edad de la planta o etapa de crecimiento del cultivo, el grupo de madurez, la susceptibilidad del cultivar y la concentración de inóculo, esta última influida por rotaciones cortas de cultivo.
Las condiciones climáticas favorables para la infección por A. solani son temperaturas superiores a 22 °C y humedad foliar durante más de 8 horas. Además, la alternancia de períodos húmedos y secos favorece el desarrollo de la enfermedad.
Un factor importante para la progresión de la enfermedad es la etapa de crecimiento del cultivo. Además de la propagación en cascada dentro de las plantas, desde las hojas más viejas hacia las más jóvenes, las plantas más viejas son significativamente más susceptibles debido al momento más temprano de emergencia tras la siembra.
La infestación de tubérculos es causada por conidios que son lavados hacia el suelo. El hongo puede ingresar al tejido del tubérculo a través de lenticelas o lesiones mecánicas en la piel durante la cosecha.
El tizón temprano también puede aparecer en otras plantas hospedantes además de la papa. Se ha observado en hospedantes como tomate (Solanum lycopersicum L.), berenjena (S. melongena L.), hierba mora vellosa (S. sarrachoides Sendt), hierba mora negra (S. nigrum L), ortiga de caballo (S. carolinense L.), pimiento (Capsicum spp.) y malezas no solanáceas (Jones et al. 1993; Pscheidt 1985; Hausladen y Aselmeyer 2017).
La diferenciación de especies de Alternaria dentro del grupo de esporas grandes o del grupo de esporas pequeñas basada en rasgos morfológicos requiere mucho tiempo y experiencia. El enfoque molecular para delimitar aislados de esporas grandes de manera rápida y precisa es un análisis multilocus basado en diferentes regiones génicas parciales, como las regiones espaciadoras transcritas internas 1 y 2 y el ADNnr 5.8S intermedio (ITS), gliceraldehído-3-fosfato deshidrogenasa (GAPDH), segunda subunidad mayor de la ARN polimerasa (RPB2), factor de elongación de la traducción 1-alfa (TEF1) y el gen del principal alérgeno de Alternaria (Alt a 1) (Woudenberg et al. 2014).
En función de las secuencias génicas de RPB2 y calmodulina, A. solani puede distinguirse de A. grandis y A. protenta (Landschoot et al. 2017a). Para las especies de esporas pequeñas, pueden utilizarse para la diferenciación la gliceraldehído-3-fosfato deshidrogenasa (gapdh), la segunda subunidad mayor de la ARN polimerasa (rpb2), el factor de elongación de la traducción 1-alfa (tef1), el gen del principal alérgeno de Alternaria (Alt a 1), la endopoligalacturonasa (endoPG), una región génica anónima (OPA10-2) y el gen de la histona H3 (Woudenberg et al. 2015; Landschoot et al. 2017a).
Impacto del tizón temprano
Después del tizón tardío de la papa (Phytophthora infestans), el tizón temprano representa hoy una de las enfermedades fúngicas más importantes de la papa. Puede encontrarse en la mayoría de los países productores de papa (Woudenberg et al. 2014), pero A. solani se describe como un patógeno muy importante especialmente en zonas más cálidas, debido a su requerimiento de temperaturas más altas.
En particular en cultivares de papa susceptibles, así como en localidades con mayor presencia de A. solani, la enfermedad puede causar pérdidas considerables de rendimiento. El daño potencial inducido por el tizón temprano en diferentes países se muestra en la tabla inferior.
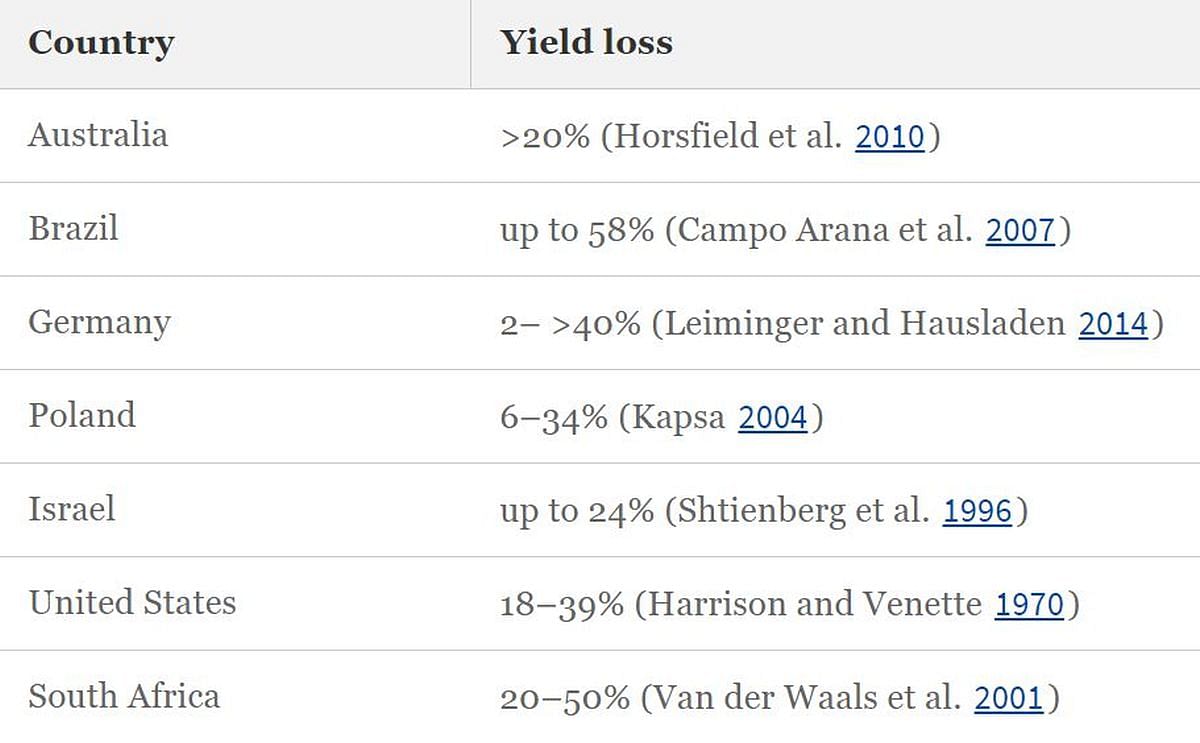
Pérdida de rendimiento debido al tizón temprano en diferentes regiones.
La necrosis del tejido foliar y la considerable defoliación causada por la infestación de Alternaria reducen el área de asimilación de la planta de papa y, por lo tanto, tienen un efecto negativo sobre el tamaño de los tubérculos y el contenido de almidón. Alternaria solani también puede infectar los tubérculos.
La posterior pudrición dura de los tubérculos conduce a una menor calidad para la comercialización como papa de mesa y para procesamiento. En Europa, la aparición de infecciones en tubérculos se conoce solo en años excepcionales.
Poblaciones del patógeno
La diversidad genética entre aislados de A. solani es relativamente alta para un hongo que se reproduce asexualmente. Además, cuando se analiza mediante ensayos de virulencia (VC), microsatélites amplificados al azar (RAMS), ADN polimórfico amplificado al azar (RAPD) y técnicas de marcadores AFLP, no se observa una agrupación clara de aislados según origen geográfico, año o incluso procedencia del mismo campo (Van der Waals et al. 2004; Leiminger et al. 2013; Odilbekov et al. 2016).
Existe polimorfismo incluso dentro del ADN mitocondrial, generalmente muy conservado. Con respecto al gen cytb, Leiminger et al. (2014) detectaron dos genotipos en poblaciones alemanas de A. solani que diferían en su estructura intrón-exón.
La presencia de los dos genotipos fue confirmada en poblaciones de A. solani en Estados Unidos (Bauske et al. 2018), Bélgica (Landschoot et al. 2017b) y Suecia (Odilbekov et al. 2016).
El registro de fungicidas específicos para Alternaria con modos de acción de sitio único para papa modificó la estructura poblacional de A. solani en muchos países debido a la aparición de mutaciones puntuales, lo que condujo a una sensibilidad reducida de los aislados mutados.
Para fungicidas inhibidores externos de la quinona (QoI), como Azoxystrobin, se ha demostrado que el cambio del aminoácido fenilalanina a leucina en la posición 129 del gen cytb (F129L) es la razón de la sensibilidad reducida en comparación con aislados de tipo silvestre (Pasche et al. 2005).
La presencia de la mutación F129L en poblaciones de A. solani se ha demostrado en diferentes países, como Estados Unidos (Pasche et al. 2004), Alemania (Leiminger et al. 2014), Suecia (Odilbekov et al. 2016) y Bélgica (Landschoot et al. 2017b).
La pérdida de sensibilidad hacia fungicidas inhibidores de la succinato deshidrogenasa (SDHI), como Boscalid, puede ser causada por diferentes mutaciones puntuales, ya que la SDH está compuesta por cuatro subunidades y el sitio de unión de los fungicidas está formado por tres de ellas (subunidades B, C y D).
Para poblaciones de A. solani, se han descrito hasta ahora cinco posibles mutaciones: H278Y y H278R en la subunidad B, H134R y H134Q en la subunidad C, y D123R y H133R en la subunidad D (Mallik et al. 2014; Metz et al. 2019).
Landschoot et al. (2017b) describen la presencia de dos mutaciones SDH diferentes en un mismo aislado. Con frecuencia, los aislados portan tanto la mutación F129L en el gen cytb como una de las mutaciones SDH (Landschoot et al. 2017b).
La sensibilidad de los aislados de referencia a fungicidas inhibidores de la desmetilación (DMI) puede variar considerablemente, pero los aislados no pertenecientes a la línea base siguen siendo sensibles, mientras que existe una pérdida clara de sensibilidad al fungicida anilinopirimidina (AP) pyrimethanil.
El modo de acción primario de las químicas AP aún no ha sido descubierto; por lo tanto, el mecanismo de resistencia es actualmente desconocido para este grupo de fungicidas ya establecido (Fonseka y Gudmestad 2016).
La resistencia contra QoI en poblaciones de A. alternata es causada por la mutación G143A en el gen cytb (Ma et al. 2003). Las mutaciones que confieren sensibilidad reducida a SDHI para esta especie de Alternaria son H277R/Y en la subunidad B de SDH, H134R en la subunidad C, y D123R y H133R en la subunidad D (Avenot et al. 2008, 2009). Fairchild et al. (2013) detectaron aislados resistentes a pyrimethanil y Fonseka y Gudmestad (2016) señalaron altos valores EC50 de DMI en aislados de referencia.
Manejo
El manejo integrado de plagas para controlar el tizón temprano requiere la implementación de varios enfoques. La enfermedad se controla principalmente mediante el uso de prácticas culturales —para reducir el inóculo presente en el suelo—, cultivares menos susceptibles y el uso de pesticidas.
Aspectos fitosanitarios
Uno de los principales componentes en este caso es la rotación de cultivos, que influye en la aparición del tizón temprano. El hongo A. solani persiste como micelio o esporas en restos vegetales o en el suelo del campo de una temporada de cultivo de papa a la siguiente.
Por lo tanto, la rotación de cultivos, incluyendo el control de plantas hospedantes como malezas —por ejemplo, hierba mora negra— en cultivos no hospedantes, reduce el inóculo inicial presente en el suelo.
Una rotación corta con cultivos hospedantes —tomate, papa— resulta en una epidemia de tizón temprano más temprana y severa (Shtienberg y Fry 1990). Además, la remoción o quema de restos vegetales infectados reduce el nivel de inóculo.
Otra opción para reducir el inóculo primario en el suelo es la biofumigación. La biofumigación consiste en la supresión de patógenos del suelo mediante isotiocianatos (ITC), que derivan de la hidrólisis de glucosinolatos por mirosinasa en células vegetales dañadas. Las plantas biofumigantes —por ejemplo, mostaza blanca y rábano forrajero— pueden reducir la progresión del tizón temprano en el cultivo (Volz et al. 2013).
Cosecha y almacenamiento de tubérculos
Alternaria solani también puede infectar el tubérculo de papa. El hongo no puede infectar a través del peridermo intacto, por lo que el riesgo de infección del tubérculo puede reducirse permitiendo que los tubérculos maduren completamente antes de la cosecha. Evitar heridas durante la cosecha y proporcionar condiciones de almacenamiento que favorezcan la cicatrización de heridas también puede reducir la infección de tubérculos (Venette y Harrison 1973).
Semilla libre de patógenos
El uso de papa semilla libre de enfermedades y virus es la base de una producción de papa económica. Las plantas de papa infectadas con virus son más susceptibles al tizón temprano que las plantas sanas.
Estrés biótico y abiótico
Las plantas de papa estresadas por factores bióticos o abióticos son más susceptibles al tizón temprano que las plantas no estresadas. Existen diferentes tipos de estrés abiótico para las plantas durante la temporada de crecimiento, como sequía, altas temperaturas y riego por aspersión.
Además, el riego por aspersión puede prolongar el período de humedad foliar, permitiendo una infección fúngica exitosa. El estrés biótico es provocado por insectos —por ejemplo, pulgones y escarabajo de la papa de Colorado—, que también son conocidos vectores de virus.
Nutrición vegetal
Una nutrición equilibrada para las plantas de papa durante el período de crecimiento es la base para un crecimiento vegetal óptimo y un buen rendimiento de papa. Una fertilidad ideal del suelo y una nutrición vegetal adecuada pueden disminuir la severidad de A. solani (Lambert et al. 2005; MacDonald et al. 2007).
Bajo condiciones específicas, como la sequía, cuando las plantas no pueden obtener suficientes nutrientes del suelo a través de las raíces, un fertilizante foliar puede reducir la deficiencia nutricional y disminuir la susceptibilidad de la planta al tizón temprano.
La forma del fertilizante también puede influir en la progresión de la enfermedad causada por A. solani. El uso de cianamida cálcica retrasa el tizón temprano, ya que los efectos secundarios fungicidas de los productos de degradación de la cianamida cálcica pueden reducir el inóculo inicial en el suelo (Volz et al. 2013).
Cultivares resistentes
Existen cultivares con susceptibilidad reducida al tizón temprano; sin embargo, hasta ahora no se han encontrado genotipos completamente resistentes. La resistencia observada en campo de las variedades frente a la infección del follaje está asociada con la madurez de la planta. En general, los cultivares de maduración temprana son más susceptibles, y los cultivares de maduración tardía son más resistentes a A. solani (Johanson y Thurston 1990; Abuley et al. 2017).
No existe correlación entre el grupo de madurez y la aparición de los primeros síntomas de tizón temprano en las hojas, pero sí una fuerte correlación entre el grupo de madurez y la progresión de la enfermedad. La epidemia en cultivares de maduración temprana comienza antes.
Resulta interesante que existen variedades dentro de un mismo grupo de madurez que son más resistentes al tizón temprano (Johanson y Thurston 1990; Leiminger y Hausladen 2014). En general, existe la posibilidad de influir en la progresión de la enfermedad mediante la plantación de cultivares más resistentes.
Uso de fungicidas
El método más común para controlar el tizón temprano en papa es el uso de pesticidas químicos. Algunos fungicidas que se utilizan para el control del tizón tardío (Phytophthora infestans) también tienen cierto efecto sobre Alternaria solani —por ejemplo, maneb, mancozeb, chlorothalonil y triphenyl tin hydroxide—. Los fungicidas más eficaces para el control del tizón temprano contienen ingredientes activos del grupo de las estrobilurinas y los azoles.
Las estrobilurinas, también conocidas como inhibidores externos de la quinona (QoI), son una clase importante de fungicidas en la agricultura porque tienen actividad de amplio espectro. Inhiben la respiración mitocondrial en hongos al unirse al sitio Qo del complejo citocromo b (cytb), bloqueando la transferencia de electrones e inhibiendo la síntesis de ATP (Bartlett et al. 2002).
Las carboxamidas (SDHI) inhiben la enzima succinato deshidrogenasa (Sdh), un componente del complejo II en la cadena de transporte de electrones mitocondrial (Kuhn 1984). Aunque los dos grupos de fungicidas tienen un modo de acción similar, los fungicidas SDHI y QoI no presentan resistencia cruzada.
El grupo de los triazoles pertenece a los fungicidas DMI, que inhiben una enzima específica, la C14-desmetilasa. Esta enzima desempeña un papel importante en la biosíntesis de ergosterol. El ergosterol es importante para la estructura funcional de la membrana y para el desarrollo de paredes celulares funcionales.
Debido al modo de acción de sitio único de las estrobilurinas (QoI) y las carboxamidas (SDHI), estos fungicidas presentan un alto riesgo de desarrollo de resistencia. En varias áreas productoras de papa se encuentran mutantes que muestran una sensibilidad reducida en ensayos in vitro e in vivo, así como una eficacia reducida en el campo (ver Poblaciones del patógeno).
La mayoría de estos fungicidas tienen una actividad curativa muy limitada y deben usarse de manera preventiva. El control del tizón temprano se logra principalmente mediante aplicaciones múltiples y frecuentes de fungicidas protectores. La optimización del uso de fungicidas para el control del tizón temprano representa un desafío considerable.
El hongo produce grandes cantidades de inóculo secundario durante la temporada de crecimiento. Por lo tanto, existen diferentes DSS —sistemas de apoyo a la toma de decisiones— disponibles para optimizar el uso de aplicaciones fungicidas en el manejo del tizón temprano en papa.
Una posibilidad es utilizar valores umbral basados en el progreso de la enfermedad (Leiminger y Hausladen 2012). En algunos países, el manejo de la enfermedad se basa en sistemas informáticos interactivos que pronostican condiciones climáticas favorables para la infección por Alternaria solani o umbrales de grados-día de temperatura.
Alternativamente, la recomendación para el uso de fungicidas se basa en el desarrollo de la planta —tamaño de la planta, edad de la planta, inicio de la floración de la papa—.
En general, la combinación de edad de la planta y resistencia del hospedante, progreso de la enfermedad y riesgo de infección calculado en función del clima se utiliza como base para un manejo integrado de plagas.
Mirando hacia el futuro
El control del tizón temprano será un desafío considerable en el futuro. La creciente importancia del tizón temprano en la papa se debe a varios factores. El cambio climático y el calentamiento global generarán condiciones más favorables para la infección, el crecimiento y el progreso de la enfermedad causada por el hongo en varias áreas productoras de papa.
El aumento de las temperaturas durante la temporada de crecimiento genera condiciones favorables para el patógeno, debido a su requerimiento de temperaturas más altas. Un mayor estrés abiótico —sequía— incrementa la susceptibilidad de la planta y, por lo tanto, también favorecerá el progreso de la enfermedad.
Además, el manejo integrado de plagas basado en tratamientos continuos con pesticidas será cada vez menos eficaz. La aparición de mutantes frente a diferentes modos de acción resultará en una menor eficiencia del control del tizón temprano mediante fungicidas.
Por lo tanto, en el futuro, el control exitoso del tizón temprano requerirá la implementación de prácticas culturales, el uso de pesticidas con atención al desarrollo de resistencia a fungicidas y el cultivo de variedades menos susceptibles.