
Introduction to Powdery Scab in Potato
Powdery scab is a major soil borne disease of potato caused by Spongospora subterranea f. sp. subterranea (Sss). The disease primarily affects potato tubers, producing raised pustules that rupture and develop into characteristic powdery lesions. These blemishes significantly reduce tuber quality and marketability in fresh, processing and seed potato markets.
Although powdery scab does not always cause substantial yield losses, it can lead to serious economic damage through tuber downgrading, rejection of seed lots, increased processing costs and reduced consumer acceptance. The disease has become increasingly important in potato-growing regions worldwide due to intensive cultivation practices, irrigation and the pathogen's exceptional ability to survive in soil for many years.
Both seed and ware potatoes are affected. In seed potato production, even low levels of infection can result in certification failures, trade restrictions and the spread of the pathogen to new areas. In commercial potato production, blemished tubers often fail to meet market quality standards, reducing profitability. Powdery scab is also of particular concern because the pathogen acts as a vector of Potato Mop-Top Virus (PMTV), a virus responsible for internal tuber necrosis and additional economic losses.
As global potato production continues to intensify, effective management of powdery scab has become essential for maintaining crop quality, protecting market access and ensuring sustainable potato production.

Characteristic lesions of powdery scab on cultivar Agria.
Causal Organism and Biology of Powdery Scab
Powdery scab is caused by Spongospora subterranea f. sp. subterranea (Sss), an obligate biotrophic pathogen belonging to the plasmodiophorids (phylum Cercozoa, class Phytomyxea). Although often described as fungus-like, it is actually a protist and differs biologically from true fungi.
The pathogen survives in soil and infected plant debris as durable resting spore aggregates known as sporosori (spore balls or cystosori). These structures can remain viable in soil for more than 10 years allowing the disease to persist even in the absence of potato crops. Under cool and moist conditions, resting spores germinate and release motile biflagellate zoospores that infect potato roots, stolons and developing tubers.
Inside host tissues, the pathogen forms multinucleate plasmodia that develop into either secondary zoosporangia for rapid multiplication or new sporosori for long-term survival. This complex life cycle enables repeated infection cycles during the growing season, particularly when soils remain wet for extended periods.
Because S. subterranea is a protist rather than a true fungus, many conventional fungicides provide limited control making integrated disease management essential.
Global Distribution and Economic Importance of Powdery Scab
Powdery scab is found in virtually all major potato producing regions worldwide and has been reported on every continent where potatoes are cultivated. The disease is particularly prevalent in temperate regions characterized by cool, moist soils, although it can also occur in warmer climates under irrigation.
Significant outbreaks have been reported throughout Europe, North America, South America, Australia, New Zealand, Asia and Africa. The pathogen is believed to have originated in the Andean region of South America, the center of origin of cultivated potato and subsequently spread internationally through infected seed tubers.
The economic importance of powdery scab extends beyond cosmetic tuber damage. In seed potato production, infections can result in certification failures and trade restrictions. In fresh and processing markets, affected tubers are often downgraded or rejected, increasing sorting, grading and peeling costs. The disease can also reduce plant vigor through root and stolon infections and contributes to the spread of Potato Mop-Top Virus (PMTV), further increasing quality losses and market risks.
With increasing global potato production and stricter quality standards, powdery scab remains one of the most challenging soil borne diseases affecting the potato industry.
Economic Consequences of Powdery Scab in Potato Production
The economic impact of powdery scab is primarily associated with reductions in tuber quality rather than direct yield losses. Lesions on potato tubers diminish their visual appeal making them unsuitable for premium fresh markets and increasing rejection rates during grading and packaging.
In processing industries, infected tubers require deeper peeling to remove scab lesions, resulting in higher processing costs and reduced recovery of usable product. Severe infections can lead to substantial financial losses when tubers fail to meet market specifications.
For seed potato producers, powdery scab represents a significant threat because infected seed lots may fail certification standards and face restrictions in domestic and international trade. The disease also increases the risk of spreading Potato Mop-Top Virus (PMTV), which can cause additional yield and quality losses through internal tuber necrosis.
Long-term management costs, including the use of certified seed, soil testing, crop rotation, drainage improvements and resistant cultivars, further contribute to the economic burden. Consequently, powdery scab remains a major constraint to profitable and sustainable potato production worldwide.
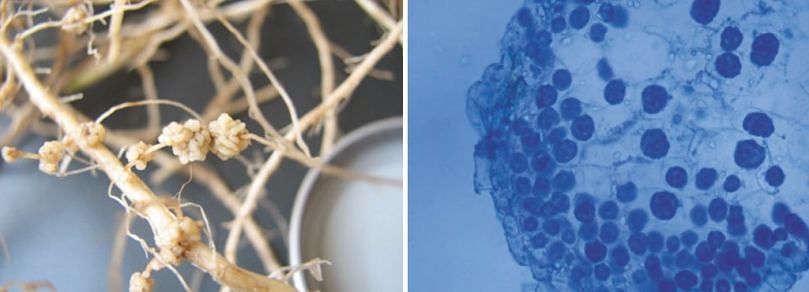
Root galling of potato caused by Spongospora subterranea (left), Sporosori inside root gall (right)
Host Range of Spongospora subterranea f. sp. subterranea
Spongospora subterranea f. sp. subterranea (Sss), the causal agent of powdery scab has a host range extending beyond its primary host, potato (Solanum tuberosum). It can infect several members of the Solanaceae family, including tomato (Solanum lycopersicum), pepper (Capsicum spp.), and common weeds such as hairy nightshade (Solanum sarrachoides), black nightshade (Solanum nigrum) and jimsonweed (Datura stramonium). These alternative hosts can support root infections and, in some cases, the production of sporosori (resting spore clusters), enabling long-term survival of the pathogen in soil. Volunteer potato plants and solanaceous weeds are particularly important reservoirs allowing the pathogen to persist between potato growing seasons.
Research has shown that S. subterranea can interact with plants across multiple families with reports of infection in more than 26 monocot and dicot families. Susceptible species include crops such as maize, onion, wheat, oat and mustard, as well as numerous weed species. Host plants are often classified according to the pathogen structures they support- non-hosts (no infection), trap crops (zoosporangial stage only), Type I hosts (sporosori production only) and Type II hosts (production of both zoosporangia and sporosori). Consequently, some rotation crops and weeds may inadvertently maintain or increase soil inoculum levels.
The broad host range complicates disease management because crop rotation alone rarely eliminates the pathogen. While certain plants may function as trap crops by supporting only limited infection without producing resting spores, true hosts capable of producing sporosori can increase pathogen populations. Effective management therefore requires careful weed control and the selection of rotation crops that minimize sporosori formation.
Disease Cycle of Powdery Scab in Potato
The disease cycle of Spongospora subterranea f. sp. subterranea is complex and polycyclic, involving both asexual (zoosporangial) and sexual/sporogenic phases. The pathogen survives in soil and infected plant debris as resting spores contained within sporosori (spore balls). Under favorable environmental conditions, these resting spores germinate and release motile, biflagellate primary zoospores. The zoospores move through water films in the soil and infect susceptible tissues, including root hairs, roots, stolons and young developing tubers through lenticels, eyes or minor wounds.
Following infection, a uninucleate plasmodium develops within host cells and subsequently becomes multinucleate through repeated nuclear divisions. During the zoosporangial phase, secondary zoospores are produced and released, resulting in repeated secondary infections and rapid pathogen multiplication during the growing season. Alternatively, during the sporogenic phase, which involves karyogamy and meiosis, the plasmodium develops into new sporosori containing numerous resting spores. As root galls and tuber lesions rupture, these sporosori are released back into the soil completing the disease cycle.
The pathogen spreads through infected seed tubers, contaminated soil adhering to machinery, equipment, footwear and vehicles, as well as irrigation water, surface runoff and manure containing viable spores. Because multiple infection cycles can occur during extended periods of favorable conditions, powdery scab epidemics may develop rapidly. This polycyclic nature explains why disease severity is often influenced more by environmental conditions than by initial inoculum levels.
")
Powdery Scab Life Cycle in Potato (Spongospora subterranea f. sp. subterranea)
Environmental Conditions Favoring Powdery Scab Disease Development
Powdery scab development is strongly favored by cool soil temperatures, typically ranging from 9–17°C with optimum conditions for zoospore release and infection occurring between 12–15°C. These temperatures enhance zoospore mobility and increase host susceptibility during critical growth stages such as stolon development and tuber initiation. Root gall formation may occur at slightly higher temperatures, whereas tuber lesion development is generally most severe under cooler conditions.
High soil moisture is essential for disease development because zoospores require free water for movement and infection. Waterlogged soils, excessive irrigation, poor drainage and prolonged periods of soil saturation significantly increase disease risk. Heavy textured clay and loam soils that retain moisture are generally more conducive to powdery scab than well drained sandy soils. Soil compaction further aggravates the problem by reducing drainage and prolonging wet conditions.
Additional factors that may contribute to disease severity include excessive nitrogen fertilization, particularly in nitrate or ammonium forms, which can stimulate vigorous root growth and potentially increase susceptibility to infection. Although powdery scab is commonly associated with cool, wet climates, the disease can also become severe in arid regions when irrigation practices create prolonged periods of soil moisture. Overall, environmental conditions often play a greater role in disease development than initial inoculum levels making proper drainage, irrigation management and field selection essential components of integrated disease management.
Recognizing Powdery Scab Symptoms in Potato
Powdery scab symptoms are primarily confined to belowground plant parts, including roots, stolons and tubers. On roots and stolons, the pathogen induces hyperplasia and hypertrophy, resulting in the formation of small, white to cream colored galls. These galls initially appear as slight swellings and gradually enlarge into irregular, warty outgrowths, typically ranging from 1–10 mm in diameter. As they mature, the galls turn brown to dark brown and eventually rupture, releasing masses of resting spores (sporosori) into the soil. Severe root and stolon infections can impair water and nutrient uptake, reduce plant vigor and contribute to yield losses under favorable disease conditions.
Tuber symptoms begin as small, purplish brown to reddish brown, slightly raised pustules or pimple like lesions, often developing around lenticels, eyes or minor wounds. As the disease progresses, these lesions enlarge into raised, wart-like swellings. The tuber skin (periderm) eventually ruptures, exposing shallow to deep cavities filled with a dark brown, powdery mass of spore balls (sporosori), which gives the disease its characteristic name. The lesions are typically surrounded by ragged remnants of the tuber skin and may vary in size and shape. Under prolonged wet conditions, lesions can expand and merge forming larger cankers that may distort tuber shape.
Symptoms often become most apparent at harvest although lesion development may continue during storage. The presence of powdery spore masses within the lesions is the key diagnostic feature distinguishing powdery scab from other tuber diseases. While tuber blemishes are the most economically important symptom because they reduce marketability and processing quality, infections of roots and stolons are critical for disease spread and inoculum buildup in the soil. Foliar symptoms are generally absent and aboveground plants may appear healthy despite significant underground infection.

Characteristic Signs and Symptoms of Potato Powdery Scab (Credit: A.J.Gevens, UW Extension)
Recognizing Powdery Scab at Different Stages
Early Stage: Initial symptoms on tubers appear as small purplish brown sunken lesions or pimple-like blisters (about 1.5 mm). These may resemble minor wounds or early infections at lenticels and eyes. On roots and stolons, early signs include small necrotic spots that begin developing into white swellings. At this stage, symptoms can be subtle and easily overlooked in the field.
Intermediate Stage: Pustules enlarge and become raised, brown and wart-like. The skin starts to crack or rupture, revealing the beginning of powdery contents. Root and stolon galls become more prominent, creamy white to tan and swollen. Lesions may show surrounding discoloration and the characteristic powdery spore masses start becoming visible upon closer inspection or gentle pressure.
Advanced/Severe Stage: Mature lesions form shallow to deeper depressions or open cankers with ragged, raised skin edges surrounding a mass of brown powdery sporosori. In severe cases, numerous lesions coalesce or cover large areas of the tuber surface leading to significant cosmetic damage and potential deformation. Root galls turn dark brown/black, enlarge significantly and begin breaking down, releasing large quantities of resting spores back into the soil. Symptoms can worsen in storage.
Accurate staging helps in timely diagnosis and management decisions. Microscopic confirmation of sporosori is recommended, especially in early or ambiguous cases.
Powdery Scab vs. Common Scab: Key Differences and Identification
Powdery scab, caused by Spongospora subterranea f. sp. subterranea and common scab, caused by several Streptomyces species are often confused because both produce blemishes on potato tubers. However, several distinctive characteristics help differentiate the two diseases.
Powdery scab lesions typically begin as small, purplish brown to reddish brown blisters that develop into raised pustules. As the lesions mature, the tuber skin ruptures, exposing a dark brown, powdery mass of resting spores (sporosori) surrounded by ragged remnants of the periderm. The lesions are generally small, round and discrete. In contrast, common scab lesions are usually corky and dry, appearing as superficial (russet), raised (erumpent) or deeply pitted scabs. These lesions are often irregular in shape and may merge to form larger scabby areas. Unlike powdery scab, common scab lesions do not contain powdery spore masses.
The two diseases also differ in their environmental preferences. Powdery scab is favored by cool (9–17°C), wet and poorly drained soils, whereas common scab is more prevalent in relatively warm, dry soils and is often associated with higher soil pH levels, typically between 5.5 and 7.5.
A key diagnostic feature of powdery scab is the presence of dark brown sporosori within ruptured lesions, which can be observed microscopically. Root and stolon galls are also characteristic of powdery scab but are not produced by common scab. Internally, powdery scab lesions are usually shallow and contain powdery spore masses, whereas common scab lesions are generally corky and may extend deeper into the tuber tissue.
Accurate identification is important because management strategies differ significantly. Powdery scab can survive in soil for many years through persistent resting spores and is also an important vector of Potato Mop-Top Virus (PMTV). Consequently, its management relies heavily on the use of disease-free seed, resistant varieties, proper drainage and moisture management. When symptoms are unclear or overlapping, laboratory confirmation through microscopy or molecular diagnostic techniques such as PCR is recommended to ensure correct disease identification and effective management.
Relationship Between Powdery Scab and Potato Mop-Top Virus (PMTV)
Spongospora subterranea f. sp. subterranea (Sss), the causal agent of powdery scab is the only known natural vector of Potato Mop-Top Virus (PMTV) making it a pathogen of major importance in potato production. PMTV is carried within the resting spores (sporosori) of S. subterranea, enabling the virus to survive in infested soils for extended periods, often exceeding 10 years and potentially much longer under favorable conditions. This prolonged persistence makes both the pathogen and the virus difficult to eliminate once established in a field.
Transmission occurs when virus carrying zoospores are released from germinating sporosori and infect potato roots, stolons or developing tubers. Following infection, the virus can spread systemically throughout the plant. PMTV is best known for causing tuber necrosis, commonly referred to as “spraing,” which appears as brown arcs, rings, lines or flecks within the tuber flesh. These internal defects significantly reduce tuber quality and marketability, particularly for fresh market and processing potatoes. Aboveground symptoms may include stunting, leaf mottling, chlorosis and distortion; however, foliar symptoms are often mild or may be completely absent making field diagnosis difficult.
Although PMTV can be transmitted through infected seed tubers, soil borne transmission via S. subterranea is considered the primary and most persistent pathway for disease spread. Importantly, potato cultivars that exhibit resistance to tuber symptoms of powdery scab may still support root infections by the pathogen allowing PMTV transmission to occur.
The close association between powdery scab and PMTV greatly increases the economic significance of S. subterranea. In addition to causing direct damage through powdery scab lesions, the pathogen facilitates the spread and long-term survival of PMTV in agricultural soils. Consequently, effective management of powdery scab is a critical component of PMTV control. In regions where both diseases occur, particularly in major potato-growing areas of Europe, North America and Australia, seed certification and phytosanitary programs emphasize the use of disease-free planting material and the avoidance of infested fields. Since there are no effective direct control measures for PMTV, management strategies focus primarily on preventing infection by controlling the vector and minimizing exposure to contaminated soils.
How Powdery Scab Is Diagnosed and Detected in Potatoes
Field Diagnosis: Field diagnosis of powdery scab is primarily based on the observation of characteristic symptoms on potato tubers, roots and stolons. Infected tubers develop raised purplish brown pustules that eventually rupture, exposing powdery masses of brown resting spores (sporosori) beneath the skin. As lesions mature, the skin breaks open, leaving irregular, corky pits with ragged edges. On roots and stolons, the disease causes the formation of white to brown galls of varying sizes. However, symptom expression can vary depending on environmental conditions and cultivar susceptibility. Early infections may be difficult to detect, symptoms can continue to develop during storage and lesions are sometimes confused with common scab caused by Streptomyces spp. Therefore, visual diagnosis alone is not always reliable. Additionally, latent infections in tubers and low levels of soil inoculum cannot be detected through field observation.
Laboratory Diagnosis: Laboratory confirmation is commonly achieved through microscopic examination of lesion scrapings, root galls or infected tissue samples. The presence of characteristic sporosori (spore balls), which consist of aggregates of thick-walled resting spores is considered a key diagnostic feature of Spongospora subterranea f. sp. subterranea (Sss). For greater sensitivity and specificity, molecular diagnostic techniques such as conventional PCR, real-time quantitative PCR (qPCR) and emerging isothermal amplification methods, including Recombinase Aided Amplification coupled with Lateral Flow Dipstick (RAA-LFD) are increasingly used. These methods can detect and quantify Sss DNA in potato tubers, roots, soil, irrigation water and other environmental samples, including asymptomatic infections. Serological assays and bait plant bioassays using susceptible potato or tomato seedlings may also be employed to assess pathogen presence and infectivity.
Soil Testing and Risk Assessment: Soil testing plays a crucial role in powdery scab management and pre-planting risk assessment. Quantitative molecular assays, particularly qPCR-based methods, enable the measurement of pathogen inoculum levels in soil helping growers identify fields with a high risk of disease development. Effective sampling involves collecting representative soil cores across the field to ensure accurate estimation of spore distribution. Testing results can guide field selection, crop rotation planning and other management decisions. Recent advances in molecular diagnostics have improved the speed, accuracy, sensitivity and field applicability of soil testing, enhancing the detection of both Spongospora subterranea f. sp. subterranea and its associated pathogen, Potato Mop-Top Virus (PMTV).
Integrated Disease Management of Powdery Scab
Disease-Free Seed and Clean Field Selection: Integrated disease management (IDM) for powdery scab emphasizes prevention and the use of multiple complementary control measures as no single strategy provides complete control due to the pathogen's long-term survival in soil. The use of certified disease-free seed tubers and planting in fields with low or no known inoculum are the foundations of disease management. Fields with a history of powdery scab, particularly poorly drained fields should be avoided whenever possible.
Crop Rotation, Weed Control and Field Sanitation: Long crop rotations of 3–10 years or more with non-host crops can help reduce pathogen populations in the soil although complete eradication is difficult because resting spores can remain viable for many years. Effective control of solanaceous weeds and volunteer potatoes is also important as these plants can serve as alternative hosts. Strict field sanitation practices, including cleaning machinery, equipment, footwear and vehicles before moving between fields help prevent the spread of contaminated soil and pathogen inoculum.
Drainage, Irrigation and Environmental Management: Powdery scab development is strongly influenced by soil moisture because the pathogen produces motile zoospores that require free water for infection. Improving soil drainage, avoiding waterlogged conditions and carefully managing irrigation during tuber initiation can reduce disease pressure. Adjusting planting dates to avoid prolonged periods of cool, wet soil conditions may further limit zoospore activity and infection.
Resistant and Tolerant Potato Varieties: The use of resistant or tolerant potato varieties is an important component of integrated disease management. Although complete resistance is not currently available, some cultivars, including many russet type varieties, exhibit reduced tuber symptoms and lower disease severity. Selecting tolerant varieties can help minimize economic losses and improve marketable yield.
Chemical Control Options: Chemical control measures provide partial suppression of powdery scab and are most effective when combined with other management practices. In-furrow or soil applied fungicides such as fluazinam, mancozeb and cyazofamid have demonstrated moderate efficacy in reducing disease severity and improving marketable yield. However, their effectiveness may vary depending on environmental conditions, soil characteristics and disease pressure.
Biological Control and Soil Amendment Strategies: Biological and soil-based approaches can provide additional disease suppression. The use of glucosinolate rich brassica cover crops, beneficial microorganisms such as Trichoderma spp. and soil amendments including boron have shown potential for reducing disease incidence or severity. These measures are generally supplementary and are most effective when integrated with other management practices.
Monitoring, Field History and Integrated Management Success: Successful long-term management of powdery scab depends on combining disease-free seed, clean field selection, crop rotation, weed control, sanitation, drainage improvement, irrigation management, tolerant varieties and appropriate chemical or biological treatments. Regular monitoring, consideration of field history and adaptation of management strategies to local environmental conditions are essential for reducing disease impact and maintaining tuber quality.
Future Research and Sustainable Management of Powdery Scab
Breeding for Durable Resistance: Future research should prioritize the development of potato varieties with durable resistance to both tuber lesions and root gall formation caused by Spongospora subterranea. Breeding efforts may benefit from marker assisted selection, genomic tools and a deeper understanding of host pathogen interactions through omics-based approaches, including genomics, transcriptomics, proteomics and metabolomics. These technologies can help identify resistance mechanisms and accelerate the development of improved cultivars.
Improved Diagnostics and Disease Detection: Advances in diagnostic technologies are needed to support early detection and precision management of powdery scab. Research is focused on developing rapid, sensitive and field deployable diagnostic tools capable of detecting the pathogen in soil, roots and seed tubers. Improved methods for the simultaneous detection of Spongospora subterranea and Potato Mop-Top Virus (PMTV) would further enhance disease monitoring and management programs.
Sustainable Soil and Crop Management Strategies: Sustainable management approaches are increasingly focused on enhancing soil health and reducing disease pressure through ecological methods. Areas of interest include soil microbiome manipulation, the identification and utilization of naturally suppressive soils and the use of elicitors to stimulate plant defense responses. Additional research is needed to optimize cultural practices such as organic amendments, crop rotations, planting date adjustments, irrigation management and other agronomic techniques that can contribute to disease suppression.
Climate Change and Disease Adaptation: Climate change may influence the future distribution and severity of powdery scab through changes in temperature, rainfall patterns and soil moisture conditions. Research is needed to model potential shifts in disease risk, identify vulnerable production regions and develop adaptive management strategies that enhance resilience under changing environmental conditions.
Pathogen Biology and Disease Interactions: Further investigation into the genetic diversity, population structure and evolution of Spongospora subterranea is essential for improving disease management and resistance breeding efforts. Understanding how the pathogen interacts with other potato diseases including secondary tuber rots and virus infections will help refine integrated disease management strategies and improve overall crop health.
Towards Sustainable and Resilient Production Systems: Long-term management goals focus on reducing reliance on chemical control measures through the development of effective biological, genetic and cultural solutions while maintaining economic sustainability for potato growers. Achieving these objectives will require strong collaboration among plant breeders, pathologists, agronomists and growers to address existing knowledge gaps and develop resilient potato production systems capable of managing powdery scab under diverse growing conditions.